
Posted on 07/05/2005 7:07:57 PM PDT by balrog666
wonderful infodump.
you know they will not read it, right?
Thank you for posting your fantasies.
and fossil records have failed to prove species change, their credibility is virtually zero.
Now you're just posting falsehoods. See the previous posts for material which refutes this nonsense. Where did you "learn" this misinformation?
Your choice of a ham-fisted cartoon to insult the intellect of those of us who do not swallow this unsubstantiated theory does little to change my mind.
It's not expected to change your closed mind. It's expected to demonstrate the ludicrousness of allowing creationists to dictate science standards, to those whose minds are open enough to understand its relevance. I think it does that rather well.
All right, you want an example? How about this?
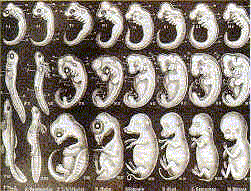
I remember this drawing, or one similar to it, which was supposed to convince my young skull full of mush that since embryos of humans, amphibians, dogs, birds, and reptiles kinda sorta looked alike, then they must all be related.
No it wasn't. You misunderstood it when you were ten, and you're misunderstanding it now.
This was published in textbooks long after it was proven to be a fake.
Not in the way you falsely claim it was, no, it wasn't. Feel free to actually quote your textbook -- or *any* textbook -- actually doing what you claim. Many creationists have made the claim (see below), but all their claims have evaporated when they were asked to actually produce these mythical textbooks. Either they slunk away into the night, never to be heard from again, or they gave the names and editions of textbooks which, when actually examined, proved to *NOT* be using the Haeckel drawings in the manner which the creationists claimed.
So go for it, maybe you can be the very first to actually document these frequent but spurious allegations of "known fakery" by the writers of biology textbooks in nefariously dishonest support of the crumbling shell of evolution, blah blah blah.
I know that that's now a popular creationist mantra, but it's a remarkably dishonest distortion of the actual situation. The details of the allegation are actually a much better example of *CREATIONIST* dishonesty and propaganda than the attempted attack on *evolutionist* honesty.
The flap started with Jonathan Wells, the author of the error-ridden anti-evolution book, "Icons of Evolution". His chapter on Haeckel's embryo drawings suffers from the same problem as all his other chapters -- it's just so jaw-droppingly wrong on so many points, and manages to twists the facts so badly (but always "concidentally" in the same direction -- in support of the author's agenda), that one is constantly wondering whether the book could be so misleading out of a) complete incompetence on the part of the author, b) complete dishonesty, or c) a combination of both.
Suffice to say, though, that Wells's presentation on the Haeckel issue is a hollow (and grossly incorrect) cartoon version of the real situation. In this respect, it is unfortunately par for the course when it comes to creationist "critiques" of evolutionary biology.
But as usual, there's a large audience for material bashing evolution, no matter how childishly flawed and transparently false, so this distortion by Wells has become vastly popular among creationists in the few short years since it was published in 2000. I see that you've uncritically swallowed it as well.
Wells's dishonest/incompetent fairy tale version of the Haeckel issue boils down to, "Haeckel faked his drawings, evolutionists (including Darwin) knew this and have used them anyway for decades in order to lie to the public, and this shows how thin/dishonest/nonexistent the evidence for evolution is, blah blah blah".
That is, in short a lie.
It's a creationist lie.
As is almost always the case whenever I look into a creationist allegation of dishonesty by evolutionists, what I found was that the truth is far more complex, interesting, and mundane than the creationists try to paint it (which again raises the usual question -- are the creationists knowingly lying, or just unable to understand anything beyond a cartoon version?), and far, *far* less a crushing blow against evolution or evolutionary biologists than the creationists would like to believe.
At the risk of oversimplifying it myself, or giving the wrong impression, the highlights of the "Cliff Notes" short version of the actual story are:
1. Contrary to Wells's misleading implication, Darwin did *not* rely in any way on Haeckel's work -- Haeckel published his work many years *after* Darwin.
2. Haeckel was a gifted artist, prolific biological researcher, and tireless popularizer of science in his time (in the last 1800's). He added much of value to the field.
3. Like many popularizers of science (or any field), he sometimes simplified his material, skipped over details, or prettied up illustrations for public presentation, striving for clarity over complete accuracy.
4. Through laziness, sloppiness, presumption, or any of a number of other possible reasons, his plates of embyronic development took this "prettying-up" process too far, and gave the impression of much more similarity at an early stage of development than is actually present. Embryos at that stage *are* more similar to each other than are embryos at a later stage of development, but not nearly as much as Haeckel's drawings indicated. A few even look as if they were just copied from each other instead of drawn from real specimens.
5. This is often taken as evidence of conscious fraud, but more prosaic explanations are possibilities as well. Haeckel may not have had actual specimens of those species at that stage of development, and drew what he actually believed they would look like, for example. Science was more casual back in those days, especially when done for public education instead of research for publication in journals. In any case, Haeckel's reasons are irrelevant to the more recent accusations being made by creationists.
6. Haeckel also had a hypothesis about evolution and embryology, usually summed up as "ontology recapitulates phylogeny" -- in short, that an animal literally retraces its past evolutionary tree as it develops from a single fertilized egg cell to a fully-developed creature ready to be born. This also probably influenced (consciously or unconsciously) what he thought he saw through his microscope and drew on paper.
7. Nonetheless, this hypothesis was examined and already discarded as incorrect by biologists by around 1900.
8. Some biologists with experience in the field of embryology also noted that Haeckel's embryo drawings were, at best, over-idealized to the point of being wrong, even in Haeckel's own time.
9. Fast-forward several decades. At some point, no one's been able to pin down exactly when, Haeckel's drawings started showing up in a few textbooks. The drawings were good-looking, textbook writers don't usually have the funds to commission all the illustrations for their textbooks (so tend to borrow heavily from existing illustrations), aren't usually top-rate experts in all the aspects of the subject they're covering, and any pre-1900 dispute in the scientific community about the accuracy of Haeckel's drawings had long since been forgotten, at least in the circles that textbook writers travel in. While experts are sometimes employed to write college-level textbooks, textbooks for gradeschool and high schools usually aren't. Haeckel's drawings of embryos are hardly the only inaccurate information to make its way into textbooks for the lower grades, but as the old saying goes, never attribute to malice that which can be adequately explained by stupidty (or carelessness, ignorance, etc.) There's no evidence that Haeckel's drawings were incorporated into textbooks with a purposeful intent to mislead students.
9. FURTHERMORE, even from the start they weren't presented in the way that Wells and other creationists falsely assert that they were -- they were not presented with the argument that "because real embryos are like the ones shown in this drawing, evolution must be true" or any such variation. Most often, they were presented in correct historical context, in order to illustrate the early -- but subsequently discarded -- notion by Haeckel that embryo development repeats evolutionary progression. The science textbook I used in school in the 1960's did exactly that. Even those textbooks which just lazily copy-and-pasted the drawings in as examples of different stages of embryos didn't make the case that evolution rested on the notion that embryos are actually as similar to each other in early development as Haeckel's drawings would indicate. Because it doesn't. It rests on the *actual* findings of embryological research (as well as findings in countless other fields) in the past 100+ years.
10. After a few decades of being a staple in various textbooks (due to some textbook writers carrying over material from earlier textbooks, etc.), and although some biologists had noticed from time to time that the illustrations weren't really accurate but hadn't made a big stink about it because textbooks generally have a *lot* of sloppy or oversimplified material in them (and doing them perfectly right would cost far more than most school districts could afford -- the classic "close enough for government work"), finally in 1997 a biologist raised the issue of Haeckel's drawings in a paper (again, as historical background) in a way that reminded a wider audience that Haeckel's drawings had some serious flaws. And that's when the creationists went nuts.
Note, however, that it was an *evolutionary biologist* who actually raised a big flag about the drawings. It wasn't a creationist "blowing the lid off" a longstanding conspiracy of evolutionist "fraud". The creationists were just the ones who decided they could misrepresent a case of inaccurate work by an early biologist and subsequent laziness by some textbook authors into some kind of Smoking Gun(tm) which allegedly demonstrated that those evil evolutionists have been knowingly lying to everyone all these years. It's horse manure, but that hasn't stopped the creationists in the past, and it wasn't going to stop them this time.
What's really funny about the creationist accusations that evolutionists have "knowingly" used Haeckel's inaccurate drawings in order to "fraudulently" make a "better" case for evolution than "actually" exists is this following observation, by the same biologist who raised the issue:
"Ironically, had Haeckel drawn the embryos accurately, his first two valid points in favor of evolution would have been better demonstrated."For more details, see:
-- Michael K. Richardson, et al., "Haeckel, Embryos, and Evolution," Science (Letters), Vol. 280 ( May 15, 1998), pp. 983-985 [emphasis mine]
Iconoclasts of Evolution: Haeckel, Behe, Wells, & the Ontogeny of a FraudWells and Haeckel's Embryos: A Review of Chapter 5 of 'Icons of Evolution'
PZ Meyers finds a textbook from 1960, and looks to see what it says about Haeckel
Paying the Price (Professor Kenneth Miller notes how Wells misrepresents Miller's textbook editions)
Abscheulich! - Atrocious! - the precursor to the theory of natural selection
If it's such ironclad, settled science,
It is.
then why all the fakery?
All *what* fakery? Your current example has fallen flat. Even if Haeckel's drawings were "fakery" at all (and they could instead be the result of laziness, etc.), they were fakes in support of a *failed* hypothesis about evolution which was discreded over a 100 years ago. They weren't fakery produced in support of Darwinian evolution or any descendant of it.
Do you -- does anyone -- have any *real* examples of actual "fakery" done in an actual attempt to falsely bolster evolutionary biology itself in the view of those who might be "on the fence"? I'm familiar with a lot more of the history of science than most people on this forum, and *I'm* not aware of any such attempts. So drop the goofy conspiracy theories, please.
Instead, Wells's accusations are examples of *creationist* fakery -- he twists the facts to give a false appearance of scientific conspiracy, where none exists.
And such lies are *incredibly* common from creationists. How about a few hundred for starters? Here's a *small* sampling of creationist dishonesty
And I'm not talking about random anonymous statements on fly-by-night websites. I'm talking about dishonest statements from leading "lights" of the creationist movement such as Henry Morris, Duane Gish, Kent Hovind, Ken Ham, Steven Austin, Carl Baugh, Jonathan Sarfati, Michael Behe, Jonathan Wells, William Dembski, Don Patton, Steve Rudd, Phillip Johnson... the list is endless.Summary of the ability of two creationists (Hovind and Havoc) to present information they *know* is false, and to *fail* to retract when reminded of their falsehoods, is presented here, along with links to all appropriate documentation.
This sort of behavior, unfortunately, is *typical* of creationists. Here, want dozens of more examples of their distortions? A few more for the road? Another? Still more, perhaps? How about even more? Ooh, here are some good examples. And there's lots more where that came from, like this< /A> and this and this and lots more here and *tons* here and countle ss more here and yet more here, a goodie... Wait, there's more over here, etc., etc., e tc., etc., etc., *ETC.* , etc., etc., etc ., . How about 300 more creationist misrepresentations? Not enough, you say? Well then visit Creationist Lies and Blunders.
Tell me, GenXFreedomFighter -- do you approve of such shameless and non-stop dishonesty from creationists?
That cartoon also tells me a lot about people who want me to believe in evolution based on the evidence to date.
You saw "lies" in your fifth-grade textbook that weren't there -- what are you "seeing" in this cartoon?
From past experience I'm pretty sure they won't, but I can always hope.
At the very least, it's always amusing to watch how they try to rationalize a truckload of actual evidence with the lies they've been told by their creationist masters who tell them that there's actually "no" evidence for evolution...
And ultimately the primary purpose is to make sure that lurkers won't mistakenly fall for the "no evidence" claptrap. Whether or not the person I'm responding to ever realizes how thoroughly they've been refuted, I trust that the lurkers won't have any problem recognizing which side has mountains of evidence supporting it -- and which side is just parroting bumper-sticker slogans with no real knowledge of the topic.

"Even a stopped clock is right twice a day.
"
only an analog clock
Uh, no they don't. Or, rather, 19th Century creationists mostly believed in fixed and separately created species, but modern creationists are invariably offended if this anachronistic view is attributed to them.
It's generally uncontroversial among creationists, for example, that all living horses and asses are probably related by common descent. (This is one of the frequently cited examples of a likely "created kind".) And note, by the way, that many of the species in this Family have widely differing chromosome numbers -- a difference not attributable solely to simple allele shifting "micro" evolution.
I wonder how GXFF found out about the internet.
Why do you feel the need to belittle and criticize the Bible by posting this banal garbage? Not everyone who believes with totality in the Bible would have such an attitude toward scientific research into disease prevention. Go back and tell your master, Satan, that he is fooling no one and his attempt to descredit the Word of God has failed.
http://www.millerandlevine.com/km/evol/embryos/Haeckel.html
When's the last time a creationist admitted and corrected an error in this manner?

Satan?
Maybe if Ich could slip in :
1. 'And Artiodactus begat Pakicetus, who begat Ambulocetus, who begat Basilosaurus.-- Cetaceans II
2. And Basilosaurus begat Odonticetus and Mysticetus.
3. Blessed was Odonticetus. And the children of Odonticetus were fruitful, and multiplied, and waxed exceeding mighty; and the waters was filled with them.'
4. However Mysticetus did not find favor in the eyes of the Lord and every man's hand was set against him. And to kill him, then render him as oil for the lamps.
(Die, spell checker, die!)
ROFL!!!
(Die, spell checker, die!)
Yeah, I'll bet it had a real field day with *that* post...
What a childish and simplistic hit piece this is. Stupid always seems to rise to the top, giving intelligent people time to start laughing at such baby boomer drivel. Might I add, liars?
That sums it up very well. I am shocked that creationsists, who claim to be Christian, defend their faith by sinning. After all, we are call upon not to bear false witness. Biologists know the limits of their knowledge regarding evolution and do not misrepresent their knowledge. Creationsist must use lies and deceit, right up to corrupting basic science, to make their flawed points. They are very poor Ambassadors for Christ and should be ashamed of themselves.
Now, it all makes sense!!!
Very good post, but the sad part is that some creationsist will come along and claim that since the inserted DNA is "broken" it is not an exact match for the viral DNA, therefore it is an assumption it came from a virus. It is really part of the original DNA of the organsim and is a coincidence that it is somewhat similar to the DNA of a virus. Since it isn't an exact match, the theory fails and there is no suppport for evolution. It was put there by God when He created the organism and it is the arrogance of Man that makes Man think it was from evolution. It's so easy to to think like these people, it is scary. No wonder such a significant proportion of the U.S. believe creationsism.
That sums it up very well. I am shocked that creationsists, who claim to be Christian, defend their faith by sinning. After all, we are call upon not to bear false witness. Biologists know the limits of their knowledge regarding evolution and do not misrepresent their knowledge. Creationsist must use lies and deceit, right up to corrupting basic science, to make their flawed points. They are very poor Ambassadors for Christ and should be ashamed of themselves.Absolutely. In fact, I will admit that the Christian faith that I had when I signed on to FR has been whittled away to agnosticism, as a direct result of the disengenuousness and sometimes outright block-headed stupidity of the creationists. If, as they say, either evolution is true or the Bible is true, the Bible is false, because evolution is not only happened, but we pretty much know how, too.
There is a very ancient thesis that the only secure foundation for public morality is religion. Moreover not just any religion will do -- it must have supernatural being(s) who monitor your behaviour, have standards of right and wrong that you must meet, and who act on how well you meet those standards.-- Evolution and Morality
We are, so to speak, the wayward children of the Gods who are to be loved, to be punished when we are bad, and rewarded when we are good.
Now the important thing for public morality is not whether there is such a God or Gods, but that people believe and accept that there are. Without such a guide, people naturally fall into depravity.
Mark that, for the purposes of securing public morality, it does not matter whether the religion is true or not, but only that people believe and accept it.
Let us accept this thesis as a postulate. Let us further accept the maintenance of public morality as a necessity, one that has higher priority than the determination of truth. Indeed, by evolutionary criteria it does. A minimum level of public morality is necessary for survival of a society; intellectual honesty is not.
Under these assumptions, what is the status of evolution? Why it is almost infinitely destructive. The funadmental objection to evolution is to a concept underlying it, the notion that it is permissable to use rational thought to study life and that, where rational thought and religion come into conflict, rational thought takes precedence.
Once such a pernicious concept takes hold, the psychology that permits the unquestioning acceptance of religious belief and its corresponding beneficial effect on behaviour is undermined, to the ultimate detriment of society. In other words:
Intellectual dishonesty in the name of religion is a virtue.
Copyright © 1993 by Richard Harter
Actually, the anti-evolution creationists aren't all baby boomers.
I'm sorry, but you all must be addressing some small fringe on the whole thing. I don't recall EVER reading in a newspaper that creationists were trying to put forth an effort to stop studying bacteria in an effort to better our health.
Just because a strain shows resistance, doesn't make it "evolved" it makes it "adaptable" to those surroundings. Proving that it can resist certain things can save lives on that fact alone. This is why even Creationists do not disapprove of these studies.
The closest I can recall hearing along these lines is Creationists battling using Zygotes for stemcell research. And that's due to 2 things: (1)Taking a human life "for science" is a very Nazi thing to do. (2) We can work with cells found in the spines of 40 year old men, or even umbilical cords, and not kill them, and still get what we need for study and saving lives.
In short: The cartoon was more insulting than informative, and definately not honest.
Of course, that may have been why it was brought to us by this particular Freeper.
Disclaimer: Opinions posted on Free Republic are those of the individual posters and do not necessarily represent the opinion of Free Republic or its management. All materials posted herein are protected by copyright law and the exemption for fair use of copyrighted works.