Posted on 02/08/2005 3:50:43 AM PST by PatrickHenry
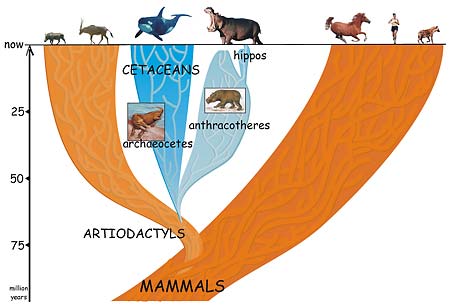
A group of four-footed mammals that flourished worldwide for 40 million years and then died out in the ice ages is the missing link between the whale and its not-so-obvious nearest relative, the hippopotamus.
The conclusion by University of California, Berkeley, post-doctoral fellow Jean-Renaud Boisserie and his French colleagues finally puts to rest the long-standing notion that the hippo is actually related to the pig or to its close relative, the South American peccary. In doing so, the finding reconciles the fossil record with the 20-year-old claim that molecular evidence points to the whale as the closest relative of the hippo.
"The problem with hippos is, if you look at the general shape of the animal it could be related to horses, as the ancient Greeks thought, or pigs, as modern scientists thought, while molecular phylogeny shows a close relationship with whales," said Boisserie. "But cetaceans – whales, porpoises and dolphins – don't look anything like hippos. There is a 40-million-year gap between fossils of early cetaceans and early hippos."
In a paper appearing this week in the Online Early Edition of the Proceedings of the National Academy of Sciences, Boisserie and colleagues Michel Brunet and Fabrice Lihoreau fill in this gap by proposing that whales and hippos had a common water-loving ancestor 50 to 60 million years ago that evolved and split into two groups: the early cetaceans, which eventually spurned land altogether and became totally aquatic; and a large and diverse group of four-legged beasts called anthracotheres. The pig-like anthracotheres, which blossomed over a 40-million-year period into at least 37 distinct genera on all continents except Oceania and South America, died out less than 2 and a half million years ago, leaving only one descendent: the hippopotamus.
This proposal places whales squarely within the large group of cloven-hoofed mammals (even-toed ungulates) known collectively as the Artiodactyla – the group that includes cows, pigs, sheep, antelopes, camels, giraffes and most of the large land animals. Rather than separating whales from the rest of the mammals, the new study supports a 1997 proposal to place the legless whales and dolphins together with the cloven-hoofed mammals in a group named Cetartiodactyla.
"Our study shows that these groups are not as unrelated as thought by morphologists," Boisserie said, referring to scientists who classify organisms based on their physical characteristics or morphology. "Cetaceans are artiodactyls, but very derived artiodactyls."
The origin of hippos has been debated vociferously for nearly 200 years, ever since the animals were rediscovered by pioneering French paleontologist Georges Cuvier and others. Their conclusion that hippos are closely related to pigs and peccaries was based primarily on their interpretation of the ridges on the molars of these species, Boisserie said.
"In this particular case, you can't really rely on the dentition, however," Boisserie said. "Teeth are the best preserved and most numerous fossils, and analysis of teeth is very important in paleontology, but they are subject to lots of environmental processes and can quickly adapt to the outside world. So, most characteristics are not dependable indications of relationships between major groups of mammals. Teeth are not as reliable as people thought."
As scientists found more fossils of early hippos and anthracotheres, a competing hypothesis roiled the waters: that hippos are descendents of the anthracotheres.
All this was thrown into disarray in 1985 when UC Berkeley's Vincent Sarich, a pioneer of the field of molecular evolution and now a professor emeritus of anthropology, analyzed blood proteins and saw a close relationship between hippos and whales. A subsequent analysis of mitochondrial, nuclear and ribosomal DNA only solidified this relationship.
Though most biologists now agree that whales and hippos are first cousins, they continue to clash over how whales and hippos are related, and where they belong within the even-toed ungulates, the artiodactyls. A major roadblock to linking whales with hippos was the lack of any fossils that appeared intermediate between the two. In fact, it was a bit embarrassing for paleontologists because the claimed link between the two would mean that one of the major radiations of mammals – the one that led to cetaceans, which represent the most successful re-adaptation to life in water – had an origin deeply nested within the artiodactyls, and that morphologists had failed to recognize it.
This new analysis finally brings the fossil evidence into accord with the molecular data, showing that whales and hippos indeed are one another's closest relatives.
"This work provides another important step for the reconciliation between molecular- and morphology-based phylogenies, and indicates new tracks for research on emergence of cetaceans," Boisserie said.
Boisserie became a hippo specialist while digging with Brunet for early human ancestors in the African republic of Chad. Most hominid fossils earlier than about 2 million years ago are found in association with hippo fossils, implying that they lived in the same biotopes and that hippos later became a source of food for our distant ancestors. Hippos first developed in Africa 16 million years ago and exploded in number around 8 million years ago, Boisserie said.
Now a post-doctoral fellow in the Human Evolution Research Center run by integrative biology professor Tim White at UC Berkeley, Boisserie decided to attempt a resolution of the conflict between the molecular data and the fossil record. New whale fossils discovered in Pakistan in 2001, some of which have limb characteristics similar to artiodactyls, drew a more certain link between whales and artiodactyls. Boisserie and his colleagues conducted a phylogenetic analysis of new and previous hippo, whale and anthracothere fossils and were able to argue persuasively that anthracotheres are the missing link between hippos and cetaceans.
While the common ancestor of cetaceans and anthracotheres probably wasn't fully aquatic, it likely lived around water, he said. And while many anthracotheres appear to have been adapted to life in water, all of the youngest fossils of anthracotheres, hippos and cetaceans are aquatic or semi-aquatic.
"Our study is the most complete to date, including lots of different taxa and a lot of new characteristics," Boisserie said. "Our results are very robust and a good alternative to our findings is still to be formulated."
Brunet is associated with the Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine at the Université de Poitiers and with the Collège de France in Paris. Lihoreau is a post-doctoral fellow in the Département de Paléontologie of the Université de N'Djaména in Chad.
The work was supported in part by the Mission Paléoanthropologique Franco-Tchadienne, which is co-directed by Brunet and Patrick Vignaud of the Université de Poitiers, and in part by funds to Boisserie from the Fondation Fyssen, the French Ministère des Affaires Etrangères and the National Science Foundation's Revealing Hominid Origins Initiative, which is co-directed by Tim White and Clark Howell of UC Berkeley.
Very interesting hypothesis.
It always struck me that some people try and limit God by saying he couldn't do this or that due to some law of physics.
My answer to that is that if God created the laws of physics, He could just as easily change them to suit his purpose whenever He wants.
One other thing (not to get off point) is that in the literal six day creation, you'll notice that God first created the plants and life BEFORE he created the sun and moon (In Genesis it is called the greater and lesser light, and were made on the ?? third or fourth day I think).
According to our known laws of physics, the sunlight would be required to sustain plant life, would it not?
It's really true that you can't tell the players apart without a scorecard.
LOL--if so, then the lack of tenure-track positions will surely lead to extinction!
I do have to go, my wolf/dog puppy is howling for me. He's a beautiful animal, will be 13 months old next week and weighs 100 lbs, his name is Cujo. If your answer to my question is that wolves and dogs are seperate species of animals then you must believe that man created a new species of animal. What a feat!
Even knowing the changing selection pressure, you don't know how a species will respond. It might just go extinct.
n.
The evolutionary formation of new biological species, usually by the division of a single species into two or more genetically distinct ones.
The question truly is, why can't we see this happening right now?
When Darwin discovered evolution, did evolution get upset and decide to stop in it's tracks?
Why would anyone want to choose a nomdeplume of such a depressing insect?
Perhaps one of the most fascinating lines of research opened up by the discovery of FOXP2 is the evolutionary origins of speech and language. “FOXP2 stands out,” says Dr Fisher. “It’s very unusual from an evolutionary point of view.”
I do not believe my ancestors were Apes. I am curious as to how we continue to adapt to gravity and our average height keeps increasing, how tall will we get?
Tell me, of any two consecutive fossils in the following list, do any differ so much from each other that anti-evolutionists wouldn't just write it off as "just adaptation", or "just microevolution"? [All of the listed specimens are actual fossils]
(Most of the above text is from The Transitional Vertebrate Fossils FAQ, and is the result of hard work by Kathleen Hunt, who deserves the credit. I've just extracted the relevant individual portions and assembled them into one direct fish-to-elephant sequence.) If you like that, here are a few hundred more.Fish to Amphibian transition: 1. Cheirolepis, (early Devonian, 400 million years ago) -- Primitive bony ray-finned fishes that gave rise to the vast majority of living fish. Heavy acanthodian-type scales, acanthodian-like skull, and big notocord.
2. Osteolepis (mid-Devonian, 390 million years ago) -- One of the earliest crossopterygian lobe-finned fishes, still sharing some characters with the lungfish (the other lobe-finned fishes). Had paired fins with a leg-like arrangement of major limb bones, capable of flexing at the "elbow", and had an early-amphibian-like skull and teeth.
3. Eusthenopteron, Sterropterygion (mid-late Devonian, 380 million years ago) -- Early rhipidistian lobe-finned fish roughly intermediate between early crossopterygian fish and the earliest amphibians. Skull very amphibian-like. Strong amphibian- like backbone. Fins very like early amphibian feet in the overall layout of the major bones, muscle attachments, and bone processes, with tetrapod-like tetrahedral humerus, and tetrapod-like elbow and knee joints. But there are no perceptible "toes", just a set of identical fin rays. Body & skull proportions rather fishlike.
4. Panderichthys, Elpistostege (mid-late Devonian, about 370 Mya) -- These "panderichthyids" are very tetrapod-like lobe-finned fish. Unlike Eusthenopteron, these fish actually look like tetrapods in overall proportions (flattened bodies, dorsally placed orbits, frontal bones! in the skull, straight tails, etc.) and have remarkably foot-like fins.
5. Obruchevichthys(middle Late Devonian, about 370 Mya -- Discovered in 1991 in Scotland, these are the earliest known tetrapod remains. The humerus is mostly tetrapod-like but retains some fish features. The discoverer, Ahlberg (1991), said: "It [the humerus] is more tetrapod-like than any fish humerus, but lacks the characteristic early tetrapod 'L-shape'...this seems to be a primitive, fish-like character....although the tibia clearly belongs to a leg, the humerus differs enough from the early tetrapod pattern to make it uncertain whether the appendage carried digits or a fin. At first sight the combination of two such extremities in the same animal seems highly unlikely on functional grounds. If, however, tetrapod limbs evolved for aquatic rather than terrestrial locomotion, as recently suggested, such a morphology might be perfectly workable."
6. Hynerpeton, Acanthostega, Ichthyostega (late Devonian, 360 Mya) -- A little later, the fin-to-foot transition was almost complete, and we have a set of early tetrapod fossils that clearly did have feet. The most complete are Ichthyostega, Acanthostega gunnari, and the newly described Hynerpeton bassetti (Daeschler et al., 1994). (There are also other genera known from more fragmentary fossils.) Hynerpeton is the earliest of these three genera (365 Ma), but is more advanced in some ways; the other two genera retained more fish- like characters longer than the Hynerpeton lineage did. Acanthostega still had internal gills, adding further support to the suggestion that unique tetrapod characters such as limbs with digits evolved first for use in water rather than for walking on land. Acanthostega also had a remarkably fish-like shoulder and forelimb. Ichthyostega was also very fishlike, retaining a fish-like finned tail, permanent lateral line system, and notochord. It turns out that Acanthostega's front foot had eight toes, and Ichthyostega's hind foot had seven toes, giving both feet the look of a short, stout flipper with many "toe rays" similar to fin rays. All you have to do to a lobe- fin to make it into a many-toed foot like this is curl it, wrapping the fin rays forward around the end of the limb. In fact, this is exactly how feet develop in larval amphibians, from a curled limb bud. Hynerpeton, in contrast, probably did not have internal gills and already had a well-developed shoulder girdle; it could elevate and retract its forelimb strongly, and it had strong muscles that attached the shoulder to the rest of the body (Daeschler et al., 1994).
7. Labyrinthodonts (eg Pholidogaster, Pteroplax) (late Dev./early Miss., 355 Mya) -- These larger amphibians still have some icthyostegid fish features, such as skull bone patterns, labyrinthine tooth dentine, presence & pattern of large palatal tusks, the fish skull hinge, pieces of gill structure between cheek & shoulder, and the vertebral structure. But they have lost several other fish features: the fin rays in the tail are gone, the vertebrae are stronger and interlocking, the nasal passage for air intake is well defined, etc.
Amphibian to Reptile transition: 8. Pholidogaster (Mississippian, about 330 Ma) -- A group of large labrinthodont amphibians, transitional between the early amphibians (the ichthyostegids, described above) and later amphibians such as rhachitomes and anthracosaurs.
9. Proterogyrinus (late Mississippian, 325 Mya) -- Classic labyrinthodont-amphibian skull and teeth, but with reptilian vertebrae, pelvis, humerus, and digits. Still has fish skull hinge. Amphibian ankle. 5-toed hand and a 2-3-4-5-3 (almost reptilian) phalangeal count.
10. Limnoscelis, Tseajaia (late Carboniferous, 300 Mya) -- Amphibians apparently derived from the early anthracosaurs, but with additional reptilian features: structure of braincase, reptilian jaw muscle, expanded neural arches.
11. Solenodonsaurus (mid-Pennsylvanian) -- An incomplete fossil, apparently between the anthracosaurs and the cotylosaurs. Loss of palatal fangs, loss of lateral line on head, etc. Still just a single sacral vertebra, though.
12. Hylonomus, Paleothyris (early Pennsylvanian) -- These are protorothyrids, very early cotylosaurs (primitive reptiles). They were quite little, lizard-sized animals with amphibian-like skulls (amphibian pineal opening, dermal bone, etc.), shoulder, pelvis, & limbs, and intermediate teeth and vertebrae. Rest of skeleton reptilian, with reptilian jaw muscle, no palatal fangs, and spool-shaped vertebral centra. Probably no eardrum yet.
13. Paleothyris (early Pennsylvanian) -- An early captorhinomorph reptile, with no temporal fenestrae at all.
14. Protoclepsydrops haplous (early Pennsylvanian) -- The earliest known synapsid reptile. Little temporal fenestra, with all surrounding bones intact. Had amphibian-type vertebrae with tiny neural processes. (reptiles had only just separated from the amphibians)
15. Clepsydrops (early Pennsylvanian) -- The second earliest known synapsid.
Reptile to Mammal transition: 16. Archaeothyris (early-mid Pennsylvanian) -- A slightly later ophiacodont. Small temporal fenestra, now with some reduced bones (supratemporal). Braincase still just loosely attached to skull. Slight hint of different tooth types. Still has some extremely primitive, amphibian/captorhinid features in the jaw, foot, and skull. Limbs, posture, etc. typically reptilian, though the ilium (major hip bone) was slightly enlarged.
17. Varanops (early Permian) -- Temporal fenestra further enlarged. Braincase floor shows first mammalian tendencies & first signs of stronger attachment to rest of skull (occiput more strongly attached). Lower jaw shows first changes in jaw musculature (slight coronoid eminence). Body narrower, deeper: vertebral column more strongly constructed. Ilium further enlarged, lower-limb musculature starts to change (prominent fourth trochanter on femur). This animal was more mobile and active. Too late to be a true ancestor, and must be a "cousin".
18. Haptodus (late Pennsylvanian) -- One of the first known sphenacodonts, showing the initiation of sphenacodont features while retaining many primitive features of the ophiacodonts. Occiput still more strongly attached to the braincase. Teeth become size-differentiated, with biggest teeth in canine region and fewer teeth overall. Stronger jaw muscles. Vertebrae parts & joints more mammalian. Neural spines on vertebrae longer. Hip strengthened by fusing to three sacral vertebrae instead of just two. Limbs very well developed.
19. Dimetrodon, Sphenacodon or a similar sphenacodont (late Pennsylvanian to early Permian, 270 Ma) -- More advanced pelycosaurs, clearly closely related to the first therapsids (next). Dimetrodon is almost definitely a "cousin" and not a direct ancestor, but as it is known from very complete fossils, it's a good model for sphenacodont anatomy. Medium-sized fenestra. Teeth further differentiated, with small incisors, two huge deep- rooted upper canines on each side, followed by smaller cheek teeth, all replaced continuously. Fully reptilian jaw hinge. Lower jaw bone made of multiple bones & with first signs of a bony prong later involved in the eardrum, but there was no eardrum yet, so these reptiles could only hear ground-borne vibrations (they did have a reptilian middle ear). Vertebrae had still longer neural spines (spectacularly so in Dimetrodon, which had a sail), and longer transverse spines for stronger locomotion muscles.
20. Biarmosuchia (late Permian) -- A therocephalian -- one of the earliest, most primitive therapsids. Several primitive, sphenacodontid features retained: jaw muscles inside the skull, platelike occiput, palatal teeth. New features: Temporal fenestra further enlarged, occupying virtually all of the cheek, with the supratemporal bone completely gone. Occipital plate slanted slightly backwards rather than forwards as in pelycosaurs, and attached still more strongly to the braincase. Upper jaw bone (maxillary) expanded to separate lacrymal from nasal bones, intermediate between early reptiles and later mammals. Still no secondary palate, but the vomer bones of the palate developed a backward extension below the palatine bones. This is the first step toward a secondary palate, and with exactly the same pattern seen in cynodonts. Canine teeth larger, dominating the dentition. Variable tooth replacement: some therocephalians (e.g Scylacosaurus) had just one canine, like mammals, and stopped replacing the canine after reaching adult size. Jaw hinge more mammalian in position and shape, jaw musculature stronger (especially the mammalian jaw muscle). The amphibian-like hinged upper jaw finally became immovable. Vertebrae still sphenacodontid-like. Radical alteration in the method of locomotion, with a much more mobile forelimb, more upright hindlimb, & more mammalian femur & pelvis. Primitive sphenacodontid humerus. The toes were approaching equal length, as in mammals, with #toe bones varying from reptilian to mammalian. The neck & tail vertebrae became distinctly different from trunk vertebrae. Probably had an eardrum in the lower jaw, by the jaw hinge.
21. Procynosuchus (latest Permian) -- The first known cynodont -- a famous group of very mammal-like therapsid reptiles, sometimes considered to be the first mammals. Probably arose from the therocephalians, judging from the distinctive secondary palate and numerous other skull characters. Enormous temporal fossae for very strong jaw muscles, formed by just one of the reptilian jaw muscles, which has now become the mammalian masseter. The large fossae is now bounded only by the thin zygomatic arch (cheekbone to you & me). Secondary palate now composed mainly of palatine bones (mammalian), rather than vomers and maxilla as in older forms; it's still only a partial bony palate (completed in life with soft tissue). Lower incisor teeth was reduced to four (per side), instead of the previous six (early mammals had three). Dentary now is 3/4 of lower jaw; the other bones are now a small complex near the jaw hinge. Jaw hinge still reptilian. Vertebral column starts to look mammalian: first two vertebrae modified for head movements, and lumbar vertebrae start to lose ribs, the first sign of functional division into thoracic and lumbar regions. Scapula beginning to change shape. Further enlargement of the ilium and reduction of the pubis in the hip. A diaphragm may have been present.
22. Dvinia [also "Permocynodon"] (latest Permian) -- Another early cynodont. First signs of teeth that are more than simple stabbing points -- cheek teeth develop a tiny cusp. The temporal fenestra increased still further. Various changes in the floor of the braincase; enlarged brain. The dentary bone was now the major bone of the lower jaw. The other jaw bones that had been present in early reptiles were reduced to a complex of smaller bones near the jaw hinge. Single occipital condyle splitting into two surfaces. The postcranial skeleton of Dvinia is virtually unknown and it is not therefore certain whether the typical features found at the next level had already evolved by this one. Metabolic rate was probably increased, at least approaching homeothermy.
23. Thrinaxodon (early Triassic) -- A more advanced "galesaurid" cynodont. Further development of several of the cynodont features seen already. Temporal fenestra still larger, larger jaw muscle attachments. Bony secondary palate almost complete. Functional division of teeth: incisors (four uppers and three lowers), canines, and then 7-9 cheek teeth with cusps for chewing. The cheek teeth were all alike, though (no premolars & molars), did not occlude together, were all single- rooted, and were replaced throughout life in alternate waves. Dentary still larger, with the little quadrate and articular bones were loosely attached. The stapes now touched the inner side of the quadrate. First sign of the mammalian jaw hinge, a ligamentous connection between the lower jaw and the squamosal bone of the skull. The occipital condyle is now two slightly separated surfaces, though not separated as far as the mammalian double condyles. Vertebral connections more mammalian, and lumbar ribs reduced. Scapula shows development of a new mammalian shoulder muscle. Ilium increased again, and all four legs fully upright, not sprawling. Tail short, as is necessary for agile quadrupedal locomotion. The whole locomotion was more agile. Number of toe bones is 2.3.4.4.3, intermediate between reptile number (2.3.4.5.4) and mammalian (2.3.3.3.3), and the "extra" toe bones were tiny. Nearly complete skeletons of these animals have been found curled up - a possible reaction to conserve heat, indicating possible endothermy? Adults and juveniles have been found together, possibly a sign of parental care. The specialization of the lumbar area (e.g. reduction of ribs) is indicative of the presence of a diaphragm, needed for higher O2 intake and homeothermy. NOTE on hearing: The eardrum had developed in the only place available for it -- the lower jaw, right near the jaw hinge, supported by a wide prong (reflected lamina) of the angular bone. These animals could now hear airborne sound, transmitted through the eardrum to two small lower jaw bones, the articular and the quadrate, which contacted the stapes in the skull, which contacted the cochlea. Rather a roundabout system and sensitive to low-frequency sound only, but better than no eardrum at all! Cynodonts developed quite loose quadrates and articulars that could vibrate freely for sound transmittal while still functioning as a jaw joint, strengthened by the mammalian jaw joint right next to it. All early mammals from the Lower Jurassic have this low-frequency ear and a double jaw joint. By the middle Jurassic, mammals lost the reptilian joint (though it still occurs briefly in embryos) and the two bones moved into the nearby middle ear, became smaller, and became much more sensitive to high-frequency sounds.
24. Cynognathus (early Triassic, 240 Ma; suspected to have existed even earlier) -- We're now at advanced cynodont level. Temporal fenestra larger. Teeth differentiating further; cheek teeth with cusps met in true occlusion for slicing up food, rate of replacement reduced, with mammalian-style tooth roots (though single roots). Dentary still larger, forming 90% of the muscle-bearing part of the lower jaw. TWO JAW JOINTS in place, mammalian and reptilian: A new bony jaw joint existed between the squamosal (skull) and the surangular bone (lower jaw), while the other jaw joint bones were reduced to a compound rod lying in a trough in the dentary, close to the middle ear. Ribs more mammalian. Scapula halfway to the mammalian condition. Limbs were held under body. There is possible evidence for fur in fossil pawprints.
25. Diademodon (early Triassic, 240 Ma; same strata as Cynognathus) -- Temporal fenestra larger still, for still stronger jaw muscles. True bony secondary palate formed exactly as in mammals, but didn't extend quite as far back. Turbinate bones possibly present in the nose (warm-blooded?). Dental changes continue: rate of tooth replacement had decreased, cheek teeth have better cusps & consistent wear facets (better occlusion). Lower jaw almost entirely dentary, with tiny articular at the hinge. Still a double jaw joint. Ribs shorten suddenly in lumbar region, probably improving diaphragm function & locomotion. Mammalian toe bones (2.3.3.3.3), with closely related species still showing variable numbers.
26. Probelesodon (mid-Triassic; South America) -- Fenestra very large, still separate from eyesocket (with postorbital bar). Secondary palate longer, but still not complete. Teeth double-rooted, as in mammals. Nares separated. Second jaw joint stronger. Lumbar ribs totally lost; thoracic ribs more mammalian, vertebral connections very mammalian. Hip & femur more mammalian.
27. Probainognathus (mid-Triassic, 239-235 Ma, Argentina) -- Larger brain with various skull changes: pineal foramen ("third eye") closes, fusion of some skull plates. Cheekbone slender, low down on the side of the eye socket. Postorbital bar still there. Additional cusps on cheek teeth. Still two jaw joints. Still had cervical ribs & lumbar ribs, but they were very short. Reptilian "costal plates" on thoracic ribs mostly lost. Mammalian #toe bones.
28. Pachygenelus, Diarthrognathus (earliest Jurassic, 209 Ma) -- These are trithelodontids. Inflation of nasal cavity, establishment of Eustachian tubes between ear and pharynx, loss of postorbital bar. Alternate replacement of mostly single- rooted teeth. This group also began to develop double tooth roots -- in Pachygenelus the single root of the cheek teeth begins to split in two at the base. Pachygenelus also has mammalian tooth enamel, and mammalian tooth occlusion. Double jaw joint, with the second joint now a dentary-squamosal (instead of surangular), fully mammalian. Incipient dentary condyle. Reptilian jaw joint still present but functioning almost entirely in hearing; postdentary bones further reduced to tiny rod of bones in jaw near middle ear; probably could hear high frequencies now. More mammalian neck vertebrae for a flexible neck. Hip more mammalian, with a very mammalian iliac blade & femur. Highly mobile, mammalian-style shoulder. Probably had coupled locomotion & breathing.
29. Sinoconodon (early Jurassic, 208 Ma) -- The next known very ancient proto-mammal. Eyesocket fully mammalian now (closed medial wall). Hindbrain expanded. Permanent cheekteeth, like mammals, but the other teeth were still replaced several times. Mammalian jaw joint stronger, with large dentary condyle fitting into a distinct fossa on the squamosal. This final refinement of the joint automatically makes this animal a true "mammal". Reptilian jaw joint still present, though tiny.
Proto-mammal to Placental Mammal transition: 30. Kuehneotherium (early Jurassic, about 205 Ma) -- A slightly later proto-mammal, sometimes considered the first known pantothere (primitive placental-type mammal). Teeth and skull like a placental mammal. The three major cusps on the upper & lower molars were rotated to form interlocking shearing triangles as in the more advanced placental mammals & marsupials. Still has a double jaw joint, though.
31. Eozostrodon, Morganucodon, Haldanodon (early Jurassic, ~205 Ma) -- A group of early proto-mammals called "morganucodonts". The restructuring of the secondary palate and the floor of the braincase had continued, and was now very mammalian. Truly mammalian teeth: the cheek teeth were finally differentiated into simple premolars and more complex molars, and teeth were replaced only once. Triangular- cusped molars. Reversal of the previous trend toward reduced incisors, with lower incisors increasing to four. Tiny remnant of the reptilian jaw joint. Once thought to be ancestral to monotremes only, but now thought to be ancestral to all three groups of modern mammals -- monotremes, marsupials, and placentals.
32. Peramus (late Jurassic, about 155 Ma) -- A "eupantothere" (more advanced placental-type mammal). The closest known relative of the placentals & marsupials. Triconodont molar has with more defined cusps. This fossil is known only from teeth, but judging from closely related eupantotheres (e.g. Amphitherium) it had finally lost the reptilian jaw joint, attaing a fully mammalian three-boned middle ear with excellent high-frequency hearing. Has only 8 cheek teeth, less than other eupantotheres and close to the 7 of the first placental mammals. Also has a large talonid on its "tribosphenic" molars, almost as large as that of the first placentals -- the first development of grinding capability.
33. Endotherium (very latest Jurassic, 147 Ma) -- An advanced eupantothere. Fully tribosphenic molars with a well- developed talonid. Known only from one specimen. From Asia; recent fossil finds in Asia suggest that the tribosphenic molar evolved there.
34. Vincelestes neuquenianus (early Cretaceous, 135 Ma) -- A probably-placental mammal with some marsupial traits, known from some nice skulls. Placental-type braincase and coiled cochlea. Its intracranial arteries & veins ran in a composite monotreme/placental pattern derived from homologous extracranial vessels in the cynodonts. (Rougier et al., 1992)
35. Kennalestes and Asioryctes (late Cretaceous, Mongolia) -- Small, slender animals; eyesocket open behind; simple ring to support eardrum; primitive placental-type brain with large olfactory bulbs; basic primitive tribosphenic tooth pattern. Canine now double rooted. Still just a trace of a non-dentary bone, the coronoid, on the otherwise all-dentary jaw. "Could have given rise to nearly all subsequent placentals." says Carroll (1988).
Placental mammal to elephant transition: 36. Protungulatum (latest Cretaceous) -- Transitional between earliest placental mammals and the condylarths (primitive, small hoofed animals). These early, simple insectivore- like small mammals had one new development: their cheek teeth had grinding surfaces instead of simple, pointed cusps. They were the first mammal herbivores. All their other features are generalized and primitive -- simple plantigrade five-toed clawed feet, all teeth present (3:1:4:3) with no gaps, all limb bones present and unfused, pointy-faced, narrow small brain, eyesocket not closed.
37. Minchenella or a similar condylarth (late Paleocene) -- Known only from lower jaws. Has a distinctive broadened shelf on the third molar.
38. Phenacolophus (late Paleocene or early Eocene) -- An early embrithopod (very early, slightly elephant-like condylarths), thought to be the stem-group of all elephants.
39. Pilgrimella (early Eocene) -- An anthracobunid (early proto-elephant condylarth), with massive molar cusps aligned in two transverse ridges.
40. Unnamed species of proto-elephant (early Eocene) -- Discovered recently in Algeria. Had slightly enlarged upper incisors (the beginnings of tusks), and various tooth reductions. Still had "normal" molars instead of the strange multi-layered molars of modern elephants. Had the high forehead and pneumatized skull bones of later elephants, and was clearly a heavy-boned, slow animal. Only one meter tall.
41. Moeritherium, Numidotherium, Barytherium (early-mid Eocene) -- A group of three similar very early elephants. It is unclear which of the three came first. Pig-sized with stout legs, broad spreading feet and flat hooves. Elephantish face with the eye set far forward & a very deep jaw. Second incisors enlarged into short tusks, in upper and lower jaws; little first incisors still present; loss of some teeth. No trunk.
42. Paleomastodon, Phiomia (early Oligocene) -- The first "mastodonts", a medium-sized animals with a trunk, long lower jaws, and short upper and lower tusks. Lost first incisors and canines. Molars still have heavy rounded cusps, with enamel bands becoming irregular. Phiomia was up to eight feet tall.
43. Gomphotherium (early Miocene) -- Basically a large edition of Phiomia, with tooth enamel bands becoming very irregular. Two long rows cusps on teeth became cross- crests when worn down. Gave rise to several families of elephant- relatives that spread all over the world. From here on the elephant lineages are known to the species level.
44a. The mastodon lineage split off here, becoming more adapted to a forest browser niche, and going through Miomastodon (Miocene) and Pliomastodon (Pliocene), to Mastodon (or "Mammut", Pleistocene).
44b. Meanwhile, the elephant lineage became still larger, adapting to a savannah/steppe grazer niche:
45. Stegotetrabelodon (late Miocene) -- One of the first of the "true" elephants, but still had two long rows of cross-crests, functional premolars, and lower tusks. Other early Miocene genera show compression of the molar cusps into plates (a modern feature ), with exactly as many plates as there were cusps. Molars start erupting from front to back, actually moving forward in the jaw throughout life.
46. Primelephas (latest Miocene) -- Short lower jaw makes it look like an elephant now. Reduction & loss of premolars. Very numerous plates on the molars, now; we're now at the modern elephants' bizarre system of one enormous multi-layered molar being functional at a time, moving forward in the jaw.
47. Primelephas gomphotheroides (mid-Pliocene) -- A later species that split into three lineages, Loxodonta, Elephas, and Mammuthus:
The Pleistocene record for elephants is very good. In general, after the earliest forms of the three modern genera appeared, they show very smooth, continuous evolution with almost half of the speciation events preserved in fossils. For instance, Carroll (1988) says: "Within the genus Elephas, species demonstrate continuous change over a period of 4.5 million years. ...the elephants provide excellent evidence of significant morphological change within species, through species within genera, and through genera within a family...."
- Loxodonta adaurora (5 Ma). Gave rise to the modern African elephant Loxodonta africana about 3.5 Ma.
- Elephas ekorensis (5 Ma), an early Asian elephant with rather primitive molars, clearly derived directly from P. gomphotheroides. Led directly to:
- Elephas recki, which sent off one side branch, E. hydrusicus, at 3.8 Ma, and then continued changing on its own until it became E. iolensis.
- Elephas maximus, the modern Asian elephant, clearly derived from
- E. hysudricus. Strikingly similar to young E. hysudricus animals. Possibly a case of neoteny (in which "new" traits are simply juvenile features retained into adulthood).
- Mammuthus meridionalis, clearly derived from P. gomphotheroides. Spread around the northern hemisphere. In Europe, led to M. armeniacus/trogontherii, and then to M. primigenius. In North America, led to M. imperator and then M. columbi.
Species-species transitions among the elephants:
- Maglio (1973) studied Pleistocene elephants closely. Overall, Maglio showed that at least 7 of the 17 Quaternary elephant species arose through smooth anagenesis transitions from their ancestors. For example, he said that Elephas recki "can be traced through a progressive series of stages...These stages pass almost imperceptibly into each other....In the late Pleistocene a more progressive elephant appears which I retain as a distinct species, E. iolensis, only as a matter of convenience. Although as a group, material referred to E. iolensis is distinct from that of E. recki, some intermediate specimens are known, and E. iolensis seems to represent a very progressive, terminal stage in the E. recki specific lineage."
- Maglio also documented very smooth transitions between three Eurasian mammoth species: Mammuthus meridionalis --> M. armeniacus (or M. trogontherii) --> M. primigenius.
- Lister (1993) reanalyzed mammoth teeth and confirmed Maglio's scheme of gradual evolution in European mammoths, and found evidence for gradual transitions in the North American mammoths too.
Similar fossil sequences can be listed for the majority of other major-group transitions.
(Did I hear a creationist in the back row say something about "no transitional fossils?")
Note that the changes between any two sequential transitionals are small enough that most creationists would write them off as only "microevolution" -- and yet those 50-or-so "microevolutionary" steps turn a fish into an elephant, which even the most stubborn creationist would have to concede is "macroevolution".
Once you've answered the first question, here's a second one for you: If evolutionary common descent *hasn't* actually happened -- if the different animal "kinds" were just *poofed* into existence fully-formed -- then why is it possible to order known fossils into such a smooth "transitional" chain *at all*, in a way that makes sense and is chronologically, morphologically, genetically, paleontologically, geographically (etc. etc.) consistent with the (allegedly) "non-existent" evolutionary transitions? And no, it's not possible to assemble a sequence of fossils in just any damned order you want, so don't try *that* excuse -- even evolutionary biologists aren't capable of putting together a transitional fossil sequence "showing", say, a cat evolving into a bird, or a butterfly into a bat. Please explain.
That life is so marvellously related to all other life is not something to be amazed at, for this priesthood--life a club to beat over the heads of the Stubbornly Superstitious.
Your worldview is relevant because it causes you to handwave away the evidence presented to you.
Many of our ancestor species were "human" but they weren't really us.
In fact he was promoting an alternative idea (life and diversity come from space) hence the ridicule. IIRC he also coined the term Big Bang to ridicule that theory, again because he had a competing one.
Ignorance is my strength placemarker.
Yes, let there be stories of discoveries without the dreary accompaniment of dogma.
An article about science is certainly a welcome thing in a contentious forum. I came across an article in Scientific American a few weeks ago that so blew my mind that I wanted to print it here--it was a pictorial explanation of how the lens in the eye works.
The eye is a fabulous thing, anyway--and before I had read about the lens I had taken it for granted as one of the less interesting features of this miraculous thing called "vision"-- cameras and processors made of flesh.
The lens is largely comprised of layers of transparent cells which are, for most intents and purposes, dead--just alive enough to continue. Yet it moves to allow for manipulating light and images (even for the nearsighted, like me)--
I didn't post it because the article was so dependent on the marvelous artwork and illustrations--but it was a thrilling article.
Wow, so much failure to address the points -- such a lame dodge! You have no substantive response, here.
From what? Evolution only works with what's available. The dinosaurs were gone, so no dinosaurs were available to take their places. Birds and mammals vied for the new niches, with the birds gaining temporary ascendency before the mammals displaced them in most terrestrial ecological niches.
Disclaimer: Opinions posted on Free Republic are those of the individual posters and do not necessarily represent the opinion of Free Republic or its management. All materials posted herein are protected by copyright law and the exemption for fair use of copyrighted works.